Характер взаимосвязи процессов клеточного деления

Между процессом накопления критической массы клетки, репликацией ДНК и построением клеточной перегородки не обнаружено облигатно-реципрокной связи, при которой подавление одного из процессов тормозило бы другие и наоборот. Например, в случае Bacillus subtitis возможно построение перегородки и формирование клеток нормального размера после подавления репликации ДНК налидиксовой кислотой. В результате одна из дочерних клеток не содержит ДНК. Кстати, такие клетки, не содержащие ДНК, нечувствительны к пенициллину, вызывающему лизис только активно растущих клеток, поэтому данный антибиотик можно использовать для получения их чистой популяции без ДНК для дальнейших исследований.
Можно получить и обратную картину, если ингибировать построение клеточной перегородки низкими концентрациями пенициллина G. Аналогично действует повышение температуры в случае некоторых л-мутантов. При этом рост клетки и репликация ДНК могут продолжаться, приводя к возникновению «многонуклеоидных» нитей, которые после удаления ингибитора фрагментируются на соответствующее число нормальных клеток.
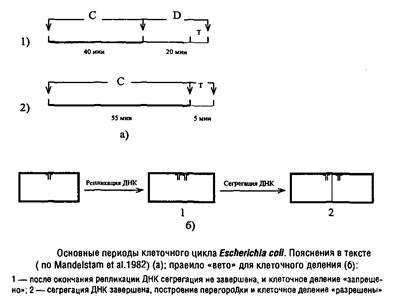
Замечено, что клеточный цикл прокариот, таких как Escherichia coli, при росте на минеральной среде с глюкозой можно подразделить на два основных периода. Они получили обозначения периодов Си D. Иногда в периоде D выделяют еще период Т – время от появления первых признаков клеточной перегородки до завершения клеточного деления.
Период С в норме занимает около 40 мин., фактически представляя собой время полной репликации генома Escherichia coli, которое мало зависит от скорости роста. В последнем случае инициация нового цикла репликации ДНК происходит до завершения клеточного деления, и дочерние клетки получают уже частично реплицированную ДНК, так что к моменту деления репликация успевает завершиться.
Период D занимает около 20 мин. – между моментом завершения репликации и моментом окончательного формирования клеточной перегородки.
Для нормального протекания клеточного цикла необходимо, чтобы в период С происходила не только репликация ДНК, но и синтез белка и РНК, так как ингибиторы транскрипции и трансляции, введенные в течение периода С, тормозят клеточное деление и увеличивают время генерации. Если же ввести эти ингибиторы на период, не превышающий 15 мин., деление клетки завершается вовремя. Очевидно, что минимальная длительность периода D может быть равна периоду Т, т.е. времени, необходимому для сборки перегородки. Эти выводы подтверждаются фактом, что данные ингибиторы, введенные в период D, не тормозят клеточное деление. Следовательно, предшественники, необходимые для построения клеточной перегородки, и другие белки, важные для завершения деления клетки, синтезируются в период С и хранятся в резерве до начала сборки перегородки.
Центральное место в проблеме регуляции клеточного деления занимает вопрос о природе сигнала, необходимого для запуска процесса сборки клеточной перегородки. Длительное время считалось, что этим сигналом является терминация репликации ДНК, однако рассмотренные нами свидетельства, указывающие на отсутствие облигатной связи между этими процессами, делают подобное заключение сомнительным.
Недавно установлено, что подавление сегрегации вновь синтезированных цепей ДНК, достигаемого в период D за счет сборки клеточной оболочки из предшественников, препятствует завершению клеточного цикла. Поэтому можно полагать, что для нормального построения клеточной перегородки от ДНК должен быть освобожден сайт, ответственный за сборку перегородки, локализованный в экваториальной части клетки и занятый ДНК сразу после завершения ее репликации. Отсюда вывод: регуляторное взаимодействие между репликацией ДНК и построением клеточной перегородки состоит в своеобразном правиле «вето» со стороны ДНК. Если нарушен процесс нормальной сегрегации реплицированной ДНК и соответствующее место в экваториальной области клетки занято, то сборка клеточной перегородки не может быть осуществлена и клеточное деление тормозится. Формально в этом случае наблюдается зависимость между репликацией ДНК и делением клетки.
Похожие материалы:
Все, что я знаю о цимбидиумах
В этой статье я собрала всю информацию о цимбидиумах, которую смогла найти и перевести из книг в нашей библиотеке. Конечно, это далеко не все, что написано об этих роскошных орхидеях, но, как говорил незабвенный Винни: "Даже немножеч ...
Коричневые пятна на молодых ростах
Молодые листочки сеянца каттлеи вылезают вот с такой коричневой расцветкой. На прежних листах такого нет. Заболевание, или природа? Первый листик такой я мазал фундазолом, но по сравнению с последующими изменений в нем никаких нет.
Ответ ...
Рост и развитие организма
Рост и развитие организма - сложные явления, результат многих метаболических процессов и размножения клеток, увеличения их размеров, процессов дифференцировки, формообразования и т.д. Этими проблемами занимаются специалисты самого разного ...