Способы сопряжения транспорта с энергией метаболизма

Для концентрирования веществ внутри клеток необходимо превращение равновесного процесса "облегченной" диффузии в одновекторный процесс "активного" транспорта. Для этого необходима затрата энергии, т.е. создание своего рода "энергетического привода" для транспорта.
Сопряжение транслокации субстрата с энергией метаболизма осуществляется двумя основными путями.
Энергия может затрачиваться на такую химическую модификацию субстрата, которая делает его неспособным взаимодействовать с переносчиком на внутренней поверхности мембраны, а также проникать через мембрану чисто диффузионным путем, что предотвращает его "утечку" из клетки.
Энергия может затрачиваться на такую модификацию переносчика, которая делает его неспособным взаимодействовать с субстратом на внутренней поверхности мембраны, что также предотвращает "утечку" субстрата из клетки.
Системы первого типа фактически осуществляют первый этапов метаболизма этих субстратов и поэтому называются системами векторного метаболизма или реакциями переноса радикалов. К ним, например, относится фосфотрансфе-разная система транспорта Сахаров и сахароспиртов, называемая также системой векторного фосфорилирования, и некоторые другие системы. Фосфотрансферазная система опосредует следующую цепь реакций.

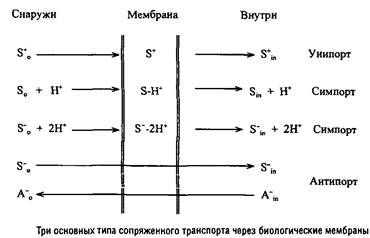
Системы второго типа, в свою очередь, подразделяются на системы "первичного" активного транспорта, генерирующие ТЭП и системы "вторичного" активного транспорта, использующие ТЭП для транспорта органических и неорганических субстратов. В некоторых случаях, например в системах со "связывающими" белками, энергия АТР непосредственно используется в транспорте субстратов. Системы "вторичного" активного транспорта распространены более широко и могут функционировать в соответствии с тремя основными механизмами.
Катионы транслоцируются в клетку по градиенту электрического потенциала путем своеобразного электрофореза.
Незаряженные соединения транслоцируются в клетку совместно с катионами Н+ или Na+.
Анионы также могут транслоцироваться в клетку путем сим-порта, присоединяя такое количество катионов, которого достаточно для перевода комплекса субстрата с переносчиком в положительно заряженную форму. Кроме того, анионы внешней среды могут обмениваться на внутриклеточные анионы. По механизму антипорта могут транслоцироваться и катионы, например у прокариот широко распространена система антипорта Н+ и Na+, а у эукариот - система антипорта К+ и Na+.
Похожие материалы:
Синергетическая парадигма в
современном естествознании
Современные физические концепции строения материи приписывают ей свойства динамизма, развития, эволюционный характер. Научному мировоззрению, по крайней мере, с XIX века была присуща идея развития
. Но после открытия Кельвином и Клаузиусо ...
Хозяйственное значение семейства Зонтичных
Зонтичные очень заметные растения, нередко они играют важную роль в сложении растительного покрова и придают своеобразные черты ландшафту. Многие зонтичные имеют большое хозяйственное значение в качестве пищевых (преимущественно овощных и ...
Способы передвижения у простейших животных. Типы органелл передвижения
Движения простейших отличаются большим разнообразием, причем в этом типе простейших животных встречаются способы локомоции, которые совершенно отсутствуют у многоклеточных животных. Это своеобразный способ передвижения амеб при помощи «пе ...