Синапсы

До последнего времени существовали две точки зрения о взаимосвязи между нейронами.
По мнению одних (Г. Гельд, 1895; К. Гольджи, 1880; С. Апати, 1897; А. Бете, 1903), использовавших материал беспозвоночных (особенно опыты Бете с крабом), нейрофибриллы - основной путь проведения нервных импульсов, а нейроцит - сгущение его цитоплазмы, имеющей трофическое значение. Было показано, что нейрофибриллы непрерывно переходят из одного нейрона в другой. Так возникла теория нейропиля (фибриллярной непрерывности, или нейрофибриллярной решетки). Применительно к высшим животным эта теория оказалась умозрительной, так как ее представителям не удалось убедить ученых в справедливости своего мнения из-за отсутствия демонстративных картин, типичных для беспозвоночных (крупные нервные клетки), что не типично для позвоночных, особенно высших (мелкие нервные клетки).
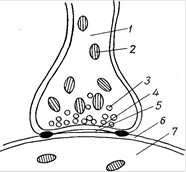
Рис.19.Схема строения синапса: 1 – пресинаптический полюс; 2 – митохондрии; 3 – светлый пресинаптический пузырек; 4 – пресинаптическая мембрана; 5 – синаптическая щель; 6 – Постсинаптическая мембрана; 7 – Постсинаптическая часть (на пре- и постсинаптической мембранах видны утолщения типа десмосом)
По мнению других (Р. Кахал, 1907; Б.И. Лаврентьев, 1939 и др.), между нейронами существует контакт - синапс (от греч. synapsis - соединение, связь). Термин дан английским физиологом Ч. Шеррингтоном. Впервые Р. Кахал установил синапсы в мозжечке: контактные связи разветвлений корзинчатых нейроцитов (вставочные нейроциты) с телами грушевидных нейроцитов (ганглиозный слой). Синапсы осуществляются передачей нервного импульса от нейрита чувствительного нейроцита к дендритам или телу корешкового (вставочного) нейроцита, а также от дендрита одного нейроцита к дендриту другого или от нейрита к нейриту двух нейроцитов. Кроме того, передача нервных импульсов может осуществляться от корешкового нейроцита к эпителиальным или мышечным клеткам.
Отсюда различают следующие синапсы: аксодендритические (контакт с дендритом последующего нейроцита), аксосоматические (контакт с телом последующего нейроцита), аксоаксональные (контакт между нейритами двух нейроцитов), дендродендритические (контакт между дендритами двух нейроцитов), аксоэпителиальные и аксомышечные (соответственно контакт корешковых нейроцитов с эпителио- и миоцитами).
Разные виды синапсов более или менее широко распространены в нервной системе. Так, аксосоматические синапсы имеют широкое распространение в нервной системе (корзинки нервных волокон на теле грушевидных нейроцитов, нейроны спинного, головного мозга, коры большого мозга, вегетативной нервной системы). Кроме того, известно, что нейрит одного нейрона, например чувствительного, может давать разветвления и оканчиваться на многих других нейронах (до 10000 синапсов). В свою очередь с одним и тем же нейроном, например корешковым нейроцитом, могут образовывать синапсы также много (1200-1800) нейронов.
Прижизненное существование синапсов у позвоночных впервые установлено в лаборатории Б.И. Лаврентьева на примере изучения внутрисердечных узлов амфибий (лягушка, жаба) с использованием метода, предложенного в 1939 г. Е.М. Граменицким (растянутая стенка предсердий на пробковом каркасе).
Прижизненными наблюдениями над синапсами и изменениями в них при тех или иных воздействиях, например ультракоротких волн, сопровождаемых возбуждением и как следствием этого желатинизацией, усилением гранулообразования (окраска янусгрюном, нейтральротом), доказано существование синапсов. Одновременно установлено совпадение наблюдаемого прижизненно и на фиксированных, а также окрашенных объектах. Субмикроскопически синапс состоит из пресинаптической, постсинаптической частей и синаптической щели. В пресинаптическую часть входит освобожденный от миелина аксон передающего нейрона, окруженный аксолеммой и содержащий в аксоплазме много митохондрий и до 3 млн. пресинаптических пузырьков диаметром 20-50 нм. В пузырьках находится медиатор, чаще всего ацетилхолин. Постсинаптическая часть представлена дендритом или телом воспринимающего нейрона, эпителиоцитом или миоцитом. Синаптическая щель расположена между пресинаптической и постсинаптической мембранами (рис. 19). Пресинаптическая мембрана - аксолемма передающего нейрона с пресинаптическим уплотнением. Элементы глии не внедряются в синаптическую щель. Постсинаптическая мембрана - цитолемма воспринимающего нейрона или другой клетки с постсинаптическим уплотнением. В ней находятся холинорецепторы (воспринимают действие ацетилхолина). Биохимическая активность в передаче нервного импульса принадлежит аксону передающего нейрона (пресинаптической части). Установлено, что перед передачей нервного импульса с нейрона на нейрон, при возбуждении передающего нейрона (пресинаптическая часть) пресинаптические пузырьки лопаются, медиатор изливается в синаптическую щель, возбуждает холинорецепторы постсинаптической мембраны и снижает ее электрический потенциал до критического. В результате этого в дендрит или тело воспринимающего нейрона (или другой клетки) усиленно проникают ионы натрия и в смежной зоне с постсинаптической мембраной возникает потенциал действия, т.е. нервный импульс. Передача нервных импульсов осуществляется всегда в одном направлении. Все это в значительной мере подтверждает справедливость взглядов представителей теории синаптических, контактных связей между нейронами. Доказано, что синаптические связи между нейронами (чувствительным, двигательным и др.) осуществляются при участии элементов нейроглии - синаптоглии (Куто, 1953). Это положение показывает, что в структуре и в функциональных проявлениях межнейронных синапсов и нервных окончаний имеются общие черты, так как и они, как правило, состоят из разветвлений аксонов, сопровождаемых нейроглией. Значение теории синапсов заключается в том, что она правильно объясняет полярность нейронов (направленность нервных импульсов от дендритов через тело к нейриту). Это положение подтверждается и данными о гистогенезе нервной ткани с установлением связи между нейронами и иннервируемыми ими тканями. Итак, нервная система едина, она состоит из нейронов и нейроглии, связанных между собой морфологически и функционально. Отсюда выдвинутая в свое время нейронная теория (В.М. Бехтерев, В. Гис, В. Вальдейер, Р. Кахал), которая трактовала, что нейрон - автономная единица нервной системы, не может быть признана вполне правильной. Рациональным в нейронной теории является то, что она подчеркивает функциональное значение нейрона в целом и в известной мере объясняет взаимоотношение центров и проводников в составе нервной системы. Накопившиеся в последующем данные позволяют внести дополнения в классическую нейронную теорию. Эти данные свидетельствуют о том, что нейрон и нейроглия взаимосвязаны, нейроны и глия очень реактивны к разнообразным внешним воздействиям и изменяются, но вместе с тем обладают значительными резервными возможностями для компенсации и восстановления нарушенных функций.
Похожие материалы:
Период цветения
Женщины всегда любили носить цветки, приколотые к одежде в специальных случаях, и, когда крупноцветковые каттлеи стали достаточно доступны, на них пал главный выбор. Каттлеи имели крупные, изумительные цветки, к тому же долгоживущие. Они ...
Горячий душ для адаптации и реанимации орхидей
Прежде чем начать разговор о горячем душе в культуре орхидей, я хочу предостеречь критически настроенных людей: если вы выработали свой собственный подход к их адаптации и реанимации – спокойно следуйте своим путем!
Не нужно губить расте ...
Деятельность эндокринных тканей в органах, обладающих неэндокринными
функциями
Эндокринные функции поджелудочной железы. Эндокринную функцию в поджелудочной железе выполняют скопления клеток эпителиального происхождения, получившие название островков Лангерганса и составляющие всего 1–2% массы поджелудочной железы. ...