Построение новой клеточной оболочки

Необходимо различать пролиферацию цитоплазматической мембраны и клеточной стенки и сегрегацию поверхностных структур.
При изучении пролиферации используют, как правило, синхронные культуры микроорганизмов и изучают включение меченных радиоизотопами соединений путем равновесного или импульсного введения этих соединений.
Таким путем установлено, что включение белков в цитоплазматическую мембрану Escherichia coli и Bacillus subtilis следует сложной кинетике, свидетельствующей о запасании предобразованных белков в цитоплазме, в период подготовки клеточного деления и быстрой их мобилизации – в процессе построения клеточной перегородки. В период деления возрастает активность некоторых литических ферментов, участвующих в образовании «брешей» в предсуществующем каркасе клеточной стенки, необходимых для включения новых ее фрагментов. Таким образом, регуляция активности этих ферментов осуществляется путем временного перевода их в скрытое состояние с последующей мобилизацией в необходимый момент. Точных данных о механизмах такой регуляции нет, но можно полагать, здесь имеет место взаимодействие ферментов с мембранами.
При изучении сегрегации поверхностных слоев также используют введение в эти структуры меченых предшественников с прослеживанием их судьбы через несколько генераций после переноса клеток на среду, не содержащую метки. Наблюдения обычно осуществляют методом электронно-микроскопической радиоавтографии, где в качестве метки используется тритий, который в силу небольшой энергии р-частиц дает на радиоавтографах короткие треки, удобные для определения мест локализации метки.
Другой подход – наблюдение за образованием и распределением маркеров структурных компонентов оболочки в течение нескольких генераций после их индукции. В этом случае удобно использовать специфические маркеры клеточной стенки или цитоплазматической мембраны, или, наконец, такие общие маркеры, как жгутики.
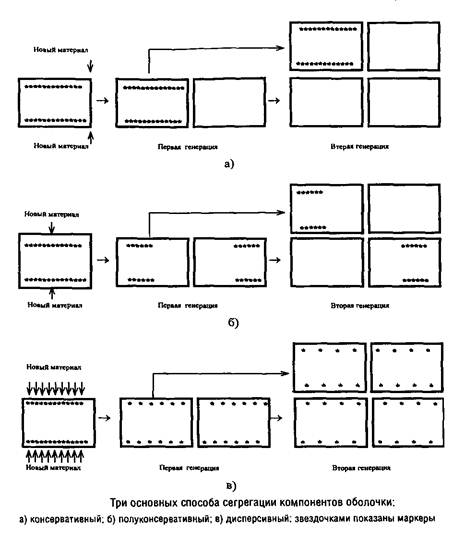
Можно представить себе три основных способа локализации сайтов включения предшественников: консервативный, полуконсервативный и дисперсивный. В первом случае после второй генерации лишь четверть клеток содержит маркеры, во втором случае – половина клеток, а в третьем – все клетки.
Вопрос о механизме сегрегации поверхностных слоев можно считать более или менее однозначно решенным лишь для кокковидных форм бактерий в случае, если они характеризуются мономорфным клеточным циклом и делятся в одной плоскости. Для этих форм разные экспериментальные подходы дают сходную картину, указывающую на полуконсервативный способ сегрегации. Для палочковидных форм бактерий сведения о способе сегрегации противоречивы.
Однозначное установление локализации мест включения мембранных компонентов затрудняется их значительной латеральной подвижностью, составляющей, например, для липополисахарида наружной мембраны Escherichia coti около 1 мкм за 25 с. Кроме того, способ сегрегации может определяться скоростью роста микроорганизма: у медленно растущих клеток Escherichia coii он близок к биполярному, а у быстро растущих становится днсперсивным.
Похожие материалы:
Однолистная-двулистная
Читал-читал про одно - двулистные каттлеи, а она (С. Penny Kuroda x C. Fitz Eugene Dixon) вон чего навыпускала. Отсюда вопрос - бывают ли они четырехлистные? Или как это объяснить. И еще - если у одной Cattleya есть и однолистные, и двули ...
Круговорот кислорода
В круговороте кислорода отчетливо выражены активная геохимическая деятельность живого вещества, его первостепенная роль в этом процессе. Биогеохимический цикл кислорода является планетарным процессом, который связывает атмосферу и гидросф ...
Теперь, давайте подытожим вышесказанное
Фаленопсисы подходят начинающим цветоводам. Им требуется особый субстрат - сосновая кора с добавкой мха, древесного угля и пластиковые горшки. Поливать их нужно так, чтобы субстрат чуть-чуть просыхал между поливами, при этом долгую просуш ...