Мембранные липидыСтраница 1

Наиболее поражает в мембранных липидах их огромное разнообразие. Причины этого пока не ясны, хотя становится все более очевидно, что, по-видимому, связано это с тем разнообразием функций, которые липиды выполняют в мембранах. Но, конечно, главная функция мембранных липидов состоит в том, что они формируют бислойный матрикс, с которым взаимодействуют белки. Основные классы липидов представлены на рис. 1.8; их краткое описание дано ниже.
Глицерофосфолипиды
Это наиболее распространенные липиды. Одна из гидроксильных групп глицерола связана с полярной группировкой, содержащей фосфат, а две другие — с гидрофобными остатками. Номенклатура гли-церидов основана на системе стереоспецифической нумерации. Если глицерол изобразить в проекции Фишера, так что центральная группа будет расположена слева, то атомы углерода будут нумероваться так, как показано на рис. В этом случае в название глицерофосфолипида вводят приставку sn для обозначения положения заместителя. В литературе встречается несколько систем стереохимических обозначений: sn, D/L и R/S. На рис. представлена стереохимия атома С-2 в этих трех системах. Природные фосфолипиды.
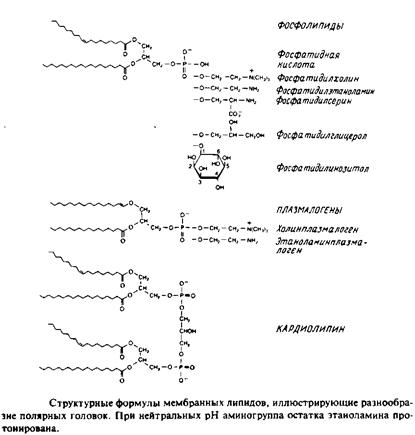
У большинства фосфоглицеридов фосфатная группа находится в ял-3-положении глицерола; она обычно связана с какой-либо из групп, представленных на рис.
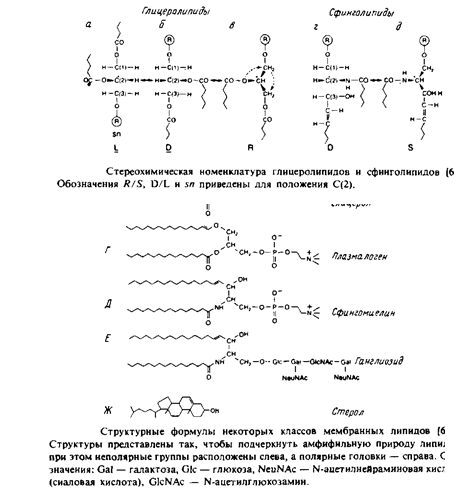
Длинные углеводородные цепи, находящиеся в положениях sn-\ и sn-2, могут присоединяться за счет сложноэфирной и простой эфирной связей. Эти цепи значительно различаются по длине, раз-ветвленности и степени ненасыщенности.
1. 1,2-диацилфосфоглицериды или фосфолипиды. Эти липиды, являющиеся сложными эфирами жирных кислот и глицерола, широко представлены во многих мембранах эукариотических и прокарио-тических клеток, за исключением архебактерий. Фосфатидил-холин является основным компонентом мембран животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы.
 Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. Степень ненасыщенности может быть разной, но чаще всего встречаются ненасыщенные кислоты 18:1, 18:2, 18:3 и 20:4. Здесь первое число обозначает длину ацильной цепи, а второе указывает на число содержащихся в ней двойных связей. Почти все природные кислоты характеризуются г/ис-конфигурацией двойных связей. Цепь в такой конфигурации имеет излом, что нарушает упаковку липидных молекул в бислое. В составе молекул многих фосфолипидов имеется одна насыщенная и одна ненасыщенная цепи. В случае животных клеток ненасыщенные цепи обычно находятся в 5и-2-положении глицерола. Такое же распределение цепей характерно и для фосфолипидов клеток Е. coli. Двойные связи в полиненасыщенных цепях обычно являются несопряженными. В фосфолипидах некоторых бактериальных мембран обнаружены разветвленные цепи, а также цепи, содержащие циклы, и гидроксильные группы в /3-положении. На рис. 1.11 показана структура некоторых из этих кислот.
Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. Степень ненасыщенности может быть разной, но чаще всего встречаются ненасыщенные кислоты 18:1, 18:2, 18:3 и 20:4. Здесь первое число обозначает длину ацильной цепи, а второе указывает на число содержащихся в ней двойных связей. Почти все природные кислоты характеризуются г/ис-конфигурацией двойных связей. Цепь в такой конфигурации имеет излом, что нарушает упаковку липидных молекул в бислое. В составе молекул многих фосфолипидов имеется одна насыщенная и одна ненасыщенная цепи. В случае животных клеток ненасыщенные цепи обычно находятся в 5и-2-положении глицерола. Такое же распределение цепей характерно и для фосфолипидов клеток Е. coli. Двойные связи в полиненасыщенных цепях обычно являются несопряженными. В фосфолипидах некоторых бактериальных мембран обнаружены разветвленные цепи, а также цепи, содержащие циклы, и гидроксильные группы в /3-положении. На рис. 1.11 показана структура некоторых из этих кислот.
2. У архебактерий глицерофосфолипиды имеют обращенную сте-реохимическую конфигурацию, при которой фосфорильные группы находятся в sn-\-положении глицерола. У многих бактерий этого вида гидрофобные компоненты представляют собой не сложные эфиры жирных кислот, а простые изопранильные эфиры глицерола.
Кардиолипины или дифосфатидилглицеролы. По сути это димерные формы фосфолипидов. Они содержатся в боль шом количестве во внутренней мембране митохондрий, в мембране хлоропластов и в некоторых бактериальных мембранах, но редко встречаются в других мембранах.
Похожие материалы:
Пути гуманизации современного высшего образования
Основой в воспитании этичного отношения к живому у учащегося наряду с родительской школой должен служить процесс гуманизации образования. Отказ от психологии пресловутого покорителя природы и замена ее сознанием ответственности человека ...
Концепция «3R»
Концепция 3R [reduction, refinement and replacement] – сокращения, усовершенствования и замены по отношению к экспериментированию на лабораторных животных была впервые предложена Расселом и Берчем [Russel&Burch] в их трактате под наз ...
Вывод
Подводя итог, надо отметить, что на сегодняшний день нельзя точно сказать, вредно или безопасно использование мобильного телефона. Исследования в данной области проводятся, но их результаты неоднозначны. Для окончательного прояснения обст ...