Фотохимический этап. Электронно-транспортная цепь фотосинтеза. Представления
о функционировании двух фотосистем. Фотофосфорилирование. Системы фотоокисления
воды и выделения кислорода при фотосинтезСтраница 2

Интересно отметить, что комплекс цитохромов Ь6 — f функционирует в системе транспорта электронов при фотосинтезе аналогично комплексу III (цит. Ь, C1) в митохондриях.
Разница в уровнях энергии между П680 и П700 (> 50 кДж) вполне достаточна для фосфорилирования AДФ, так как величина высокоэнергетической фосфатной связи АТФ равна 30,6 кДж/моль (7,3 ккал). Перепады энергии в ФС I еще значительнее. Однако само по себе освобождение энергии при нециклическом и циклическом транспорте электронов не объясняет, каким образом осуществляется фотофосфорилирование.
Механизм фосфорилирования AДФ, сопряженного с деятельностью ЭТЦ , объясняет хемиосмотическая теория, разработанная английским биохимиком П. Митчеллом (1961 — 1966). Для объяснения процессов фотофосфорилирования эта теория была впервые использована А. Ягендорфом (1967).
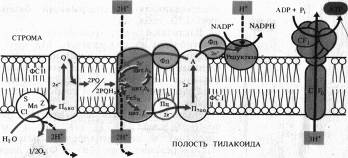
Сущность хемиосмотической теории состоит в следующем. Цепь переносчиков электронов и прогонов, действующая в соответствии с окислительно-восстановительным градиентом, перешнуровывает мембрану таким образом, что трансмембранный перенос е- и Н+ в одну сторону чередуется с переносом в обратную сторону только е-. В результате функционирования такого механизма (Н + -помпы) по одну сторону мембраны накапливается избыток Н+ и возникает электрохимический (т. е. электрический и концентрационный) потенциал ионов Н+, который служит формой запасания энергии. Обратный пассивный ток ионов Н+ через протонный канал Н+-АТФазы, получивший название сопрягающего фактор CF1, сопровождается образованием высокоэнергетической фосфатной связи АТФ.
На рис. 6.1 представлена упрощенная схема расположения переносчиков в мембране тилакоидов, объясняющая механизм фотофосфорилирования. На внутренней стороне мембраны тилакоида под действием света возбуждаются П680 и П700. Электроны от П680 захватываются акцепторами е- на наружной стороне и передаются на окисленную форму пластохинона (PQ) — липидорастворимого переносчика прогонов и электронов, выполняющего функцию челнока. Принимая два электрона, 2PQ диффундируют к цитохромному комплексу и захватывают из стромы 2Н + . Электроны с 2PQH2 поступают в цепь: F.eSR -> цит. f -> Пц,-> П700, а ионы Н + попадают в полость тилакоида. Вторая пара ионов Н+ освобождается там же при фотоокислении воды. В ФС I электроны П700 захватываются акцептором А, передаются ферредоксину и затем НAДФ+ на наружной стороне мембраны, причем 1Н+ используется на восстановление НAДФ.
Таким образом, из среды, окружающей тилакоид, при поглощении пигментами квантов света исчезают, а во внутренней полости тилакоида появляются протоны. В результате на мембране возникает электрохимический потенциал ионов Н + , который затем используется для фосфорилирования AДФ. Этот процесс называется нециклическим фотофосфорилированием.
При циклическом фотофосфорилировании, когда функционирует только ФС I, электроны от Фд поступают на цитохромный комплекс с использованием пула PQ, который действует как переносчик электронов и протонов. Затем электроны через цитохром f и Пц возвращаются на основной энергетический уровень в П700, а протоны поступают в полость тилакоида.
Похожие материалы:
Тресковые.
Минтай, атлантическая треска и другие рыбы из семейства тресковых дают около 15% мирового улова рыбы, занимая второе место после сельдеобразных.
Атлантическая треска
– одна из самых крупных рыб в семействе тресковых: до 180 см в длину. ...
Плечевая кость
Плечевая кость – длинная, трубчатая кость, состоит из диафиза и двух эпифизов. На ней различают краниальную, каудальную, медиальную и латеральную поверхности. На проксимальном эпифизе у лося находится крупная, округлая головка, сдвинутая ...
Орхидеи для начинающих.
Еще недавно орхидеи считались цветами для состоятельных людей или коллекционеров, но в последнее время ситуация изменилась. В Великобритании, например, в настоящее время они занимают одно из трех первых мест в рейтинге самых популярных ко ...