Фотохимический этап. Электронно-транспортная цепь фотосинтеза. Представления
о функционировании двух фотосистем. Фотофосфорилирование. Системы фотоокисления
воды и выделения кислорода при фотосинтезСтраница 3

Белковые комплексы, участвующие в фотохимических реакциях фотосинтеза, в мембранах хлоропластов распределены неравномерно. Наибольшие различия наблюдаются между теми мембранами, которые плотно состыкованы друг с другом в гранах и теми участками мембран, которые контактируют со стромой (закругленные участки тилакоидов гран и мембраны тилакоидов стромы).
Светособирающий комплекс и пигмент-белковый комплекс ФС II находятся в основном в мембранах, плотно контактирующих друг с другом, причем ССК играет особую роль. В адгезии тилакоидных мембран.
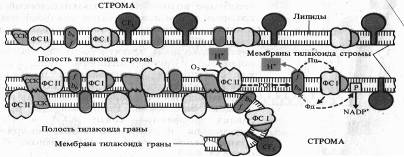
Показано, что у мутанта хламидомонады, лишенного белка, с которым связан хлорофилл Ь,
граны не образуются. ФС I со своим светособирающим комплексом преимущественно, а АТФазный комплекс (CF1 + CF0), как правило, находятся на несостыкованных участках мембран. Комплекс цитохромов b6 — f равномерно распределен как в состыкованных, так и в не состыкованных мембранах. Такое распределение белковых комплексов, участвующих в световой фазе фотосинтеза, ставит вопрос о способе их взаимодействия. Показано, что это взаимодействие осуществляется с помощью легкоподвижного липофильного пластохинона PQ
в липидной фазе и благодаря перемещению водорастворимого пластоцианина вдоль внутренних поверхностей ламелл и водорастворимого ферредоксина вдоль их
наружных поверхностей.
К латеральным перемещениям вдоль мембраны способны и сами белковые комплексы. На это передвижение влияет величина их электрического заряда. Например, восстановление пластохинона PQ
фотосистемой II приводит к активации киназы и фосфорилированию ССКа-ь.
Фосфорилирование ССК увеличивает его отрицательный заряд, что способствует перемещению ССК в стромальную область мембраны и возрастанию миграции поглощенной энергии света к ФС I. Возросшая фотохимическая активность ФС I усиливает окисление PQ,
что, в свою очередь, приводит к инактивации киназы, фосфатаза же дефосфорилирует ССК. Таким образом, обратимое фосфорилирование ССК представляет собой петлю обратной связи в системе взаимной регуляции активности ФС I и ФС II. Механизм такой регуляции включает латеральный транспорт белковых комплексов в мембранах хлоропластов.
Похожие материалы:
Пространство – время живого в биосфере
Пространство и время – две основные формы движущейся материи – в трудах В.И. Вернадского являлись стратегической целью, некоторой «Сверхзадачей» всей его натурфилософии. Глубина проникновения в строение окружающей действительности и широк ...
Особенности кровоснабжения гипоталамуса
Ядра гипоталамуса получают обильное кровоснабжение. Капиллярная сеть гипоталамуса по своей разветвлённости в несколько раз больше, чем в других отделах ЦНС. Одной из особенностей капилляров гипоталамуса является их высокая проницаемость, ...
Определение молекулярной массы нативного белка с
помощью гидродинамических методов
Применение этих методов для мембранных белков может быть сопряжено с большими трудностями, вызванными связыванием детергента.
Для нахождения молекулярной массы белка используют два метода.
1. Прямо измеряют количество связанного детерге ...