Архитектоника коры больших полушарий мозга

Учение о структурных особенностях строения коры называется архитектоникой.Клетки коры больших полушарий менее специализированы, чем нейроны других отделов мозга; тем не менее определенные их группы анатомически и физиологически тесно связаны с теми или иными специализированными отделами мозга.
Микроскопическое строение коры головного мозга неодинаково в разных ее отделах. Эти морфологические различия коры позволили выделить отдельные корковые цитоархитектонические поля. Имеется несколько вариантов классификаций корковых полей. Большинство исследователей выделяет 50 цитоархитектонических полей, Микроскопическое строение их довольно сложное.
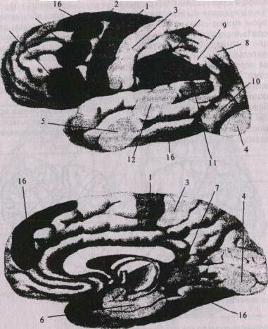
Рис. 3. Схема локализации функций в коре больших полушарий:
1 – проекционная двигательная зона; 2 – центр поворота глаз и головы в противоположную сторону; 3 – проекционная зона чувствительности; 4 – проекционная зрительная зона. Проекционные гностические зоны: 5 – слуха; 6 – обоняния; 7 – вкуса; 8 – гностическая зона схемы тела; 9 – зона стереогноза; 10 – гностическая зрительная зона; 11 – гностическая зона чтения; 12 – гностическая речевая зона; 13 – зона праксиса; 14 – праксическая речевая зона; 15 – праксическая зона письма; 16 – зона контроля за функцией мозжечка
Кора состоит из 6 слоев клеток и их волокон. Основной тип строения коры шестислойной, однако, он не везде однороден. Существуют участки коры, где один из слоев выражен значительно, а другой – слабо. В других областях коры намечается подразделение некоторых слоев на подслои и т.д.
Установлено, что области коры, связанные с определенной функцией, имеют сходное строение. Участки коры, которые близки у животных и человека по своему функциональному значению имеют определенное сходство в строении. Те участки мозга, которые выполняют чисто человеческие функции (речь), имеются только в коре человека, а у животных, даже у обезьян, отсутствуют.
Морфологическая и функциональная неоднородность коры головного мозга позволила выделить центры зрения, слуха, обоняния и т.д., которые имеют свою определенную локализацию. Однако неверно говорить о корковом центре как о строго ограниченной группе нейронов. Специализация участков коры формируется в процессе жизнедеятельности. В раннем детском возрасте функциональные зоны коры перекрывают друг друга, поэтому их границы расплывчаты и нечетки. Только в процессе обучения, накопления собственного опыта практической деятельности происходит постепенная концентрация функциональных зон в отделенные друг от друга центры.
Белое вещество больших полушарий состоит из нервных проводников. В соответствии с анатомическими и функциональными особенностями волокна белого вещества делят на ассоциативные, комиссуральные и проекционные (см. рис. 4).

Рис. 4. Нервные проводники головного мозга (схема):
а – ассоциативные волокна; б – комиссуральные волокна; в-проекционные волокна.
Ассоциативные волокна
объединяют различные участки коры внутри одного полушария. Эти волокна бывают короткие и длинные. Короткие волокна обычно имеют дугообразную форму и соединяют соседние извилины. Длинные волокна соединяют отдаленные участки коры. Комиссуальными принято называть те волокна, которые соединяют топографически идентичные участки правого и левого полушарий. Комиссуральные волокна образуют три спайки: переднюю белую спайку, спайку свода, мозолистое тело. Передняя белая спайка соединяет обонятельные области правого и левого полушарий. Спайка свода соединяет между собой гиппокамповые извилины правого и левого полушарий. Основная же масса коммисуальных волокон проходит через мозолистое тело, соединяя между собой симметричные участки обоих полушарий головного мозга.
Проекционными называют те волокна, которые связывают полушария головного мозга с нижележащими отделами мозга – стволом и спинным мозгом. В составе проекционных волокон проходят проводящие пути, несущие афферентную (чувствительную) и эфферентную (двигательную) информацию.
Итак, основные отделы, борозды и извилины головного мозга представлены на рис. 5.6.
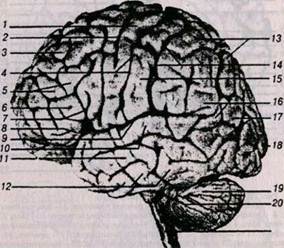
Рис. 5. Головной мозг, левое полушарие (вид сбоку):
1 – прецентральная извилина; 2 – прецентральная борозда; 3 – верхняя лобная извилина; 4 – центральная борозда; 5 – средняя лобная извилина;
б – нижняя лобная извилина; 7 – восходящая ветвь латеральной борозды;
8 – горизонтальная ветвь латеральной борозды; 9 – задняя ветвь латеральной борозды; 10 – верхняя височная извилина; 11 – средняя височная извилина;
12 – нижняя височная извилина; 13 – теменная долька; 14 – пост-центральная борозда; 15 – постцентральная извилина; 16 – надкраевая извилина;
17 – угловая извилина; 18 – затылочная доля; 19 – мозжечок; 20 – горизонтальная щель мозжечка; 21 – продолговатый мозг
Похожие материалы:
Витамин В17
Ø Химическое название: летрил, лаетраль, амигдалин.
Ø Роль в обмене веществ: обладает некоторыми противораковыми свойствами.
Ø Авитаминоз характеризуется повышенной восприимчивостью к раку, быстрой утомляемость.
&O ...
Воспаление
Воспаление - это реакция организма на внедрение инфицирующего агента, введение антигена или физическое повреждение тканей. Помимо усиления клеточной миграции, описанного выше, воспаление вызывает приток различных растворимых молекул из пл ...
Особенности строения кожных покровов и их производных с учетом различных условий
жизни хордовых животных. Особенности строения и
образования чешуи
Чешуя рыбы всегда является производным собственно кожи (кориума), и только иногда, кроме кориума, в ее образовании принимает второстепенное участие и эпидермис. Различают четыре основных типа рыбьей чешуи: плакоидную, космоидную, ганоидну ...