Нервные окончанияСтраница 1

Конечные разветвления нейронов образуют нервные окончания, которые контактируют с тканями (органами), воспринимающими нервные импульсы из центра, либо с тканями (органами), передающими воздействие внутренней и внешней среды и способствующими образованию нервных импульсов, направленных в центр. В связи с этим различают эффекторы и рецепторы.
Эффекторы, или двигательные и секреторные нервные окончания, проводят нервные импульсы от мозга к рабочему органу (мышца, железа) и регулируют его функцию, а рецепторы, или чувствительные нервные окончания, передают нервные импульсы, вызванные различными раздражителями извне (кожа, скелетная мускулатура, внутренние органы).
Эффекторы - двигательные и секреторные нервные окончания - имеют различное строение. На миоцитах неисчерченной мышечной ткани (сарколемма) они представлены в виде небольших концевых пуговок, из сплетения нейрофибрилл в виде петли, в центре которой находится скопление аксоплазмы. Субмикроскопически это окончание имеет структуру синапса (рис. 13) с типичными особенностями (см. ниже). Очень характерно, что одно нервное волокно, подходя к миоциту, делится на ветви, образующие подобие сплетения и оканчивающиеся концевыми пуговками на сарколемме. Этим увеличивается контакт нейрона с миоцитом. В месте контакта, как уже доказано, нервное окончание вырабатывает особые продукты обмена, называемые медиаторами (холин - и адренергические вещества), которые способствуют передаче импульса на рабочий орган (ткань, клетка), а также и сами могут быть причиной возбуждения и специфической работы.
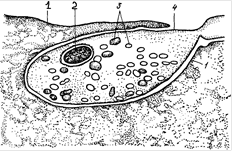
Рис. 13. Схема субмикроскопического строения двигательного нервного окончания на миоците: 1 – постсиналтнческая мембрана миоцита; 2 – митохондрия; 3 – светлые пресинаптические пузырьки; 4 – пресинаптическая мембрана нейрита
Это проверяется введением их в кровь, лимфу, когда медиаторы проявляют свое действие на другие органы (ткани).
В связи с отмеченным, полагают, что увеличение степени разветвления одного нервного волокна для образования контакта с одной мышечной клеткой способствует выработке большого количества медиатора и лучшей передаче нервного импульса.
Методом перерезки при последующей дегенерации нервных волокон, о чем будет сказано позже, доказано, что неисчерченная мускулатура иннервируется вегетативной нервной системой (постганглионарные нервные волокна).
На сердечных миоцитах (мышечные клетки миокарда сердца) в основном наблюдаются такие же нервные окончания, как и на миоцитах неисчерченной мышечной ткани. Источником иннервации также является вегетативная нервная система, причем трудно отличить окончания постганглионарных симпатических и парасимпатических волокон.
Окончания на неисчерченной и сердечной исчерченной мышечной ткани являются экстрацеллюлярными (внеклеточными), а по утверждению Ж. Буке (1913), они проникают в саркоплазму миоцитов.
Скелетная мускулатура получает иннервацию от мозга, который посылает нервные волокна к мышечным волокнам.
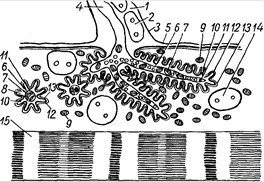
Рис. 14. Схема субмикроскопического строения нейромышечного окончания: 1 – цитоплазма нейролеммоцита; 2 – ядро нейролеммоцита; 3 – нейролемма, переходящая в наружный слой сарколеммы; 4 – аксон; 5 – сарколемма; 6 – концевые веточки нервного волокна в продольном и поперечном сечениях; 7 – митохондрии в аскоплазме: 8 – первичное синаптическое пространство; 9 – саркосомы; 10 – вторичное синаптическое пространство; 11 – светлые пресинаптические пузырьки; 12 – пресинаптическая мембрана; 13 – постсинаптическая мембрана; 14 – ядро мышечного волокна; 15 – миофибрилла, состоящая из миофиламентов
Как показывает световой микроскоп, в месте контакта с мышечным волокном нервное волокно теряет миелиновую оболочку, эндоневрий сливается с сарколеммой, которая расщепляется на два листка и включает разветвление аксона в виде петелек и пластинок из нейрофибрилл и ядер нейролеммоцитов. Окончания на мышечных волокнах скелетной мускулатуры называются нейромышечными (концевые моторные - бляшками), которые заложены внутрь сарколеммы и изолированы своим основанием - подошвой - от саркоплазмы (М.Д. Лавдовский, 1884-1885; К.А. Лавров, 1941). Ноэль (1932) доказал, что митохондрии подошвы концевой моторной бляшки близки по структуре к наблюдаемой в нейролеммоцитах и резко разнятся от митохондрий исчерченного мышечного волокна. Кроме того, при некоторых патологических процессах отмечались (Л.И. Фалин, 1954; Е.К. Плечкова, 1955) реактивные изменения в ядрах нейролеммоцитов (размножение, вакуолизация) подошвы концевых моторных бляшек, а мышечные волокна оставались без видимых изменений.
Похожие материалы:
Физико-географическая характеристика хозяйства. Рельеф,
гидрография, климат, почвы
Рельеф. Вся центральная часть хозяйства расположена на средневысоких горах. Долинные ландшафты приурочены к рекам Олекма, Нюкжа, вся остальная территория - цепь горных хребтов и плоскогорий. С севера и северо-востока подходят отроги Стано ...
Полив и субстрат
Большинство ошибок в комнатной культуре ванд связано именно с этими аспектами. Вандам, выращиваемым на свету, требуется гораздо больше воды, чем принято считать.
Для оптимального роста большинству растений этого рода необходимо чередован ...
Жидкость на росте
Сегодня обнаружил, что в молодом чехле скапливаетя вода (от автоопрыскивания) и уже там она какая то липкая или склизкая, я по возможности её удалил но чувствую, что не всю. Опасно? Загнить может, или не страшно?
Ответ:
На молодых роста ...